Cladística
| Parte de uma série sobre |
| Teoria da Evolução |
|---|
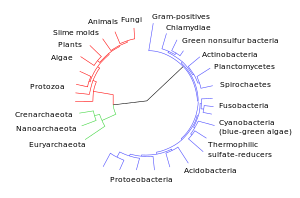 Diagrama da divergência dos grupos taxonómicos modernos em relação aos seus ancestrais comuns. |
Tópicos fundamentais
|
Processos
|
História natural
|
História da teoria da evolução
|
Campos e aplicações
|
Implicações sociais
|
|
A Cladística (gr.cl. κλάδος= ramo), também conhecida como sistemática filogenética, é uma escola da Sistemática Biológica baseada no princípio filogenético e que agrupa espécies ou táxons em grupos naturais (gerando classificações hierárquicas ou não) de acordo, unicamente, com hipóteses de relações evolutivas [1]. Segundo os fundamentos da cladística, formulados em grande parte pelo entomólogo Willi Hennig, uma classificação deve sempre expressar as relações evolutiva das espécies, não importando se as espécies são semelhantes ou diferem drasticamente entre si. Segundo esta escola, em uma classificação cada grupo deve ser obrigatoriamente Monofilético, ou seja, possuir todos os descendentes do ancestral comum mais recente do grupo estudado e nada mais. De modo que grupos parafiléticos e polifiléticos não são permitidos na classificação.[2]
O método hennigiano é explícito e diretamente conectado à ideia de que o mundo natural ordena-se de forma hierárquica, resultante do processo evolutivo. Não obstante, as classificações filogenéticas são sistemas que permitem não só o armazenamento da informação biológica, levantada para a sua construção, como também a recuperação dessa informação - os grupos monofiléticos representam padrões hierárquicos que carregam consigo a informação sobre a evolução das características das linhagens biológicas consideradas, o que não ocorre nos grupos parafiléticos e polifiléticos, dos quais pode ser depreendida apenas parte da informação usada na construção da classificação. O monofiletismo é fundamental para a sistemática filogenética e é ele que garante o sucesso do método no que tange à identificação da afinidade natural (evolutiva) entre grupos biológicos.[3]
Índice
1 Histórico
2 Princípios fundamentais da Cladística
2.1 Cladogramas
2.2 Apomorfias e plesiomorfias
2.3 Inferindo a "polaridade" de um caracter
2.4 Grupos fósseis
2.5 Critério da objetividade
3 Ver também
4 Referências
5 Ligações externas
Histórico |

Willi Hennig (1972)
Foi o entomólogo alemão Willi Hennig, nascido em Dürrhennersdorf a 20 de Abril de 1913, que desenvolveu as ideias que levariam à criação da Cladística, apresentando-as de forma sumária na sua obra Grundzüge einer Theorie der Phylogenetischen Systematik (Hennig, 1950).[4]
Usada pelos investigadores há mais de 50 anos, somente nos últimos vinte anos tornou-se popular e utilizada de forma quase universal. Este atraso na difusão das ideias básicas da cladística foi causado principalmente pelas dificuldades em torno da língua alemã, que tinha pouca representatividade no cenário científico da época, gerando um intervalo de 15 anos entre a criação e o reconhecimento da Sistemática Filogenética, e que só foi quebrado em 1966, com a publicação de “Phylogenetic Systematics”, uma nova versão do livro de 1950, feita por Willi Hennig e contando uma escrita de leitura mais simples e de mais fácil assimilação. Redigido originalmente em alemão, e traduzida posteriormente para o inglês, foi a partir desta obra que o trabalho de Hennig passou a ganhar maiores dimensões. Principalmente nos últimos 10 anos onde pode se notar um vertiginoso aumento no número de trabalhos e livros enfocados na sistemática filogenética, bem como a grande aceitação de sua estrutura teórica por novas gerações de biólogos, indicando a grande influência que este método possui.
Embora a teoria da evolução não seja obra de Hennig. Foi devido ao seu método de análise das relações de parentesco baseada em padrões evolutivos que os métodos mais antigos e laboriosos, baseados apenas em reuniões de semelhanças em vez de conhecimentos sobre os processos evolutivos, puderam ser repensados.
A cladística de Hennig forneceu os meios para integrar a enorme quantidade de conhecimento descritivo sobre os organismos, gerando uma visão unificada sobre a diversidade biológica. Zoologia e botânica, sob o enfoque filogenético, ganharam um dinamismo jamais visto antes. Sobre essa abordagem novos elementos passaram a ser incorporados em discussões sobre evolução e o conhecimento filogenético passou a ser um elemento integrador na formação dos profissionais das áreas biológicas.[5]
| “ | Fazendo da sistemática filogenética o sistema geral de referência (…) há a inestimável vantagem de que as relações com todos os outros sistemas biológicos concebíveis podem ser mais facilmente representadas através dela. Isso ocorre porque o desenvolvimento histórico dos organismos deve necessariamente estar refletido de alguma forma em todas as relações entre os organismos. Conseqüentemente, relações diretas estendem-se da sistemática filogenética para todos os outros possíveis sistemas, enquanto freqüentemente não há tais relações diretas entre esses outros sistemas.[6] | ” |
Princípios fundamentais da Cladística |
A Cladística se assenta no princípio fundamental de que os organismos devem ser classificados de acordo com as suas relações evolutivas, ou seja, as espécies devem ser classificadas de acordo com o quão recente é o seu ancestral comum. De forma que duas espécies que possuem um ancestral comum mais recente serão agrupadas em um mesmo grupo, de grau mais baixo do que o de duas espécies que possuem um ancestral comum mais distante. Quanto mais distante for o ancestral comum a duas espécies, maior será a distância entre o os seus respectivos táxons. Este processo é feito utilizando tanto espécies vivas quanto fósseis de forma continua, até que todas as espécies estejam contidas em um único grupo, contendo todos os descendentes do mais distante ancestral comum à vida.[1]
| “ | Provavelmente, tem grande significado histórico o fato de o próprio Darwin ter dito que a possibilidade de ordenar os organismos em um sistema hierárquico só é explicável assumindo-se uma relação filogenética entre eles: o simples fato de que as espécies, tanto extintas como viventes, agrupam–se em gêneros e famílias, ordens etc. – uma divisão análoga a variabilidade subjacente- não seria explicável de outra maneira, e pode não nos parecer extraordinário apenas por ser um lugar comum.[6] | ” |
O principio mais importante da Cladistica é que todos os táxons de uma filogenia devem ser monofiléticos. Partindo deste pressuposto os táxons, para a cladistica, jamais podem ser inventados, pois correspondem a entidades históricas que foram, ou ainda precisam ser, descobertas. Toda a estrutura de uma classificação deve refletir de maneira precisa e inequívoca o conhecimento disponível sobre as relações de parentesco entre os táxons incluídos na classificação. Ou seja, deve ser possível para qualquer leitor, apenas utilizando a classificação, recuperar a informação sobre as supostas relações de parentesco entre os táxons. Embora não haja uma única maneira de criar classificações estritamente filogenéticas, o fato de que as classificações devam representar inequivocamente as relações de parentesco entre os táxons é um fato incontestável para qualquer filogeneticista.[5]
Cladogramas |

Dois exemplos de cladogramas. Onde (A+B) formam um clado e (A+B+C) formam outro clado, que contém o clado (A+B)
O resultado final de uma análise cladística é obtidos na forma de árvore ou cladograma, um dendograma que expressa hipóteses de relações filogenéticas entre táxons de determinado grupo. Uma análise cladística pode ser baseada em tantas informações quanto o investigador quiser utilizar. A sistemática moderna é geralmente baseada em uma variedade de informações, incluindo seqüências de DNA (os famosos "dados moleculares"), dados bioquímicos e dados morfológicos.
Em um cladograma todos os organismos são colocados sobre as extremidades terminais dos ramos, e cada nó interior é idealmente binário, gerando dois táxons terminais. Os dois táxons de cada ramo são chamados taxa irmãs ou grupos irmão. Cada subárvore, independentemente de quantos elementos ela contenha, é chamada clado. Todos os organismos de um grupo natural estão contidos em um clado que compartilha um ancestral comum (não compartilhado com nenhum outro organismo do diagrama). Cada subtipo é definido em um número de características que aparecem nos seus membros, mas não em outras formas que não divergiu. Estas características de identificação do clado são chamados sinapomorfias (caracteres derivados compartilhados). Por exemplo, as asas anteriores endurecidas(élitros) são uma sinapomorfia de besouros.[5]
Apomorfias e plesiomorfias |

A pata pentadáctila é um estado ancestral de todos os tetrápodes
A principal característica da cladística é que esta utiliza apenas caracteres apomórficos em suas análises (caracteres derivados e "diferentes da condição ancestral"). Na terminologia cladística quando um caracter derivado é compartilhado por mais de um grupo ele é denominado Sinapomofia (do Grego syn = junto) e caracteres derivados que ocorrem em um único grupo são denominado autapomorfias e geralmente não são utilizados para determinar a relação filogenética dos grupos. Os organismos vivos também possuem Caracteres ancestrais, caracteres primitivos que herdaram sem modificações de seus ancestrais. E que são denominados plesiomorfias. Plesiomorfias que são compartilhadas por mais de um grupo de organismos são chamadas de Simplesiomorfia (sim possui o mesmo significado que o radical sin, ou seja, "em conjunto").[7]
Segundo o método de classificação cladístico, estados primitivos de caracteres, ou plesiomorfia, não podem ser usados para definir táxons. Pois não são confiáveis para determinar relações filogenéticas dentro dos grupos. Por exemplo; Se um sistemata estiver inferindo a filogenia de cão, cavalo e rã e escolhesse como caráter para classifica-los o número de dígitos dos apêndices locomotores. Utilizando este caracter a rã e o cão ficam mais aparentadas estre sí na filogenia, com cinco dígitos em cada apêndice, do que com o cavalo, com apenas um. O caráter "apêndices com cinco dígitos" é comum a todos os tetrápodes (anfíbios, répteis, aves e mamíferos), mas foi perdido nos cavalos, que reduziram seu número de dígitos de cinco para um. De forma que compartilhar o caracter "apêndices com cinco dígitos" não evidencia que cães e rãs possuem um ancestral comum mais recente do que cada um deles tem com um cavalo. Na verdade o cão e o cavalo são mamíferos e possuem um ancestral comum mais recente do que cada um deles tem com a rã, um anfíbio. Isso ilustra claramente como um caracter primitivo não é confiável para definir relações filogenéticas dentro de grupos. Ao contrário das plesiomorfias, as apomorfias são indicadores de relações filogenéticas confiáveis, por se tratarem de características que evoluíram dentro do grupo de espécies sob estudo, após o ancestral comum. Por exemplo se no exemplo anterior fosse utilizado Rã, cavalo e peixe, a pata pentadactila não seria uma condição ancestral, pois não estaria no ancestral comum aos três grupos. Ela evoluiu dentro do grupo que esta sendo analisado e informa algo sobre a filogenia. Ela informa que o cão e a rã possuem um ancestral comum mais recente do que cada um deles tem com o peixe.[1]
Inferindo a "polaridade" de um caracter |
Uma das dificuldades da análise cladística é identificar qual é efetivamente o estado de um carácter, ou a sua "polaridade" (se ele é plesiomorfico ou apomorfico). Este impasse pode ser resolvida de duas formas principais: inferindo diretamente, por dados paleontológicos sobre o estado primitivo, ou usando na análise "um grupo externo" ("outgroup"), um grupo próximo ao grupo estudado, mas ainda assim claramente distinto dele. Por exemplo; Os amniotas são um grupo conhecido particularmente por possuirem uma membrana proveniente do ovo, o âmnion, durante o seu desenvolvimento. Os amniotas são um grupo monofilético, ou seja, todos os elementos do grupo compartilham um ancestral comum. Se um investigador quisesse saber se a condição oviparidade é acestral ou derivada dentro do grupo dos amniotas. O métodos do grupo externo o permitiria concluir isto utilizando um grupo que seja parente próximo dos amniotas, mas não é amniota. Um bom exemplo seriam as salamandras, rãs e peixes, que são parentes próximos dos amniotas, mas não são amniotas. Quase todas estas espécies externas ao grupo são ovíparas.Logo, a conclusão que se obtém utilizando o método do grupo externo é a de que a oviparidade é ancestral em amniotas.
O Registro fóssil e a comparação com o grupo externo, não são as únicas formas de determinar a "polaridade" de um caracter. Existem também várias outras técnicas, como por exemplo; técnicas que utilizam o desenvolvimento embrionário, técnicas que utilizam genes parálagos e etc.. mas sem dúvida, estas são as técnicas mais utilizadas para definir a polaridade de caracteres, atualmente.[1][2]
Grupos fósseis |

Fóssil em âmbar de um Coleóptero.
Há algumas décadas, a maioria dos filogeneticista, entre eles o próprio Willi Hennig, não acreditavam que fosse possível reunir dados que permitissem concluir satisfatoriamente que uma espécie extinta fosse, de fato, ancestral. Atualmente,ao menos alguns autores admitem, que de posse de uma reconstrução filogenética, dados acumulados sobre fósseis em estratos geológicos e uma reconstrução biogeográfica dos grupos envolvidos, talvez fosse possível apresentar uma hipótese consistente indicando que uma determinada espécie extinta seja ancestral de um grupo monofilético conhecido. Ainda que se admitam hipóteses desse tipo (que seriam refutadas com a primeira autapomorfia encontrada na espécie extinta),é evidente que são necessários muitos dados acurados para a elaboração deste tipo de construção. Por outro lado é importante notar que o acúmulo de informações bastante detalhadas de fósseis em âmbar do cretáceo e do terciário pode mudar um pouco este quadro. Várias peças em âmbar permitem verificar não apenas detalhes da morfologia (e, portanto posicionar estas espécies com mais segurança em filogenias com grupos recentes),como também dados indiretos de comportamento e da biologia destes indivíduos (itens alimentares, parasitas, comensais, transporte de larvas em formigas etc). Isso conferiu um posicionamento filogenético muito mais seguro e causou uma revolução na paleontologia tradicional.[5]
| “ | A ideia de evolução orgânica foi mudada [por meio da sistemática filogenética] de uma narrativa sobre a história da vida para uma teoria científica sobre como a natureza está ordenada em uma estrutura hierárquica particular. Também ficou claro que fósseis, por muito tempo considerados a prova de que a evolução havia ocorrido, eram meramente partes dessa hierarquia, sujeitos aos mesmos limites de interpretação que os organismos vivos. Essa realização marcou o início do fim da paleontologia tradicional e de sua busca por ancestrais. A paleontologia torna-se [...] não mais a guarda de profundas verdades evolutivas, mas sim a guarda de partes extintas da hierarquia da natureza.[8] | ” |
Critério da objetividade |
Uma forma de definir qual é o melhor método de classificação é testando se ele é objetivo ou não. Uma classificação objetiva é a que representa uma propriedade natural real e livre de ambiguidades. A classificação objetiva pode ser comparada com uma classificação subjetiva. Representada por alguma propriedade escolhida arbitrariamente pelo taxonomista. Por exemplo, um taxonomista poderia decidir agrupar as espécies de acordo com o dia da semana em que ele as descobriu, algumas na segunda, outras na terça e etc... Este é um exemplo de um critério de classificação subjetivo; ele não possui embasamento teórico para justificar a sua utilização em vez da de outro. Um critério subjetivo não apresenta nenhuma propriedade inerente e exclusiva apenas aos indivíduos do grupo. Nenhum característica dos indivíduos da segunda-feira, é suficiente para distingui-los dos indivíduos da terça, quarta,quinta e sexta-feiras. Não há nada nestes indivíduos que permita a diferentes pessoas racionais, trabalhando de forma independente, chegar ao mesmo resultado que o taxonomista que as agrupou.
A cladistica tem a vantagem da objetividade. A hierarquia filogenética existe independente dos métodos que usamos para descobri-la. E é única e inequívoca quanto á forma. Quando há discordância entre diferentes técnicas para inferir relações filogenéticas, sempre há uma fonte externa ao qual o sistemata pode recorrer. Se não for possível para um sistemata descobrir a filogenia de um ou outro grupo com os dados que ele possui, ainda sim é possível assumir que tal relação exista, mesmo que ele ainda não a tenha descoberto. Com o uso de sistemas de classificação subjetivos, tal afirmativa não é verdadeira. Não existe nenhuma hierarquia natural e única, análoga á hierarquia filogenética.[1]
Ver também |
Livro: Evolução |
- Clado
- Árvore filogenética
- Homologia
- Sistemática
- Willi Hennig
Referências
↑ abcde Ridley, Mark. Evolução (3ª edição),Porto Alegre: Artmed, 2006
↑ ab Futuyma, D.J. Evolutionary Biology (2º edição), Sinauer Associates, Sunderland, Massachusetts, 1986
↑ RIEPPEL, O. Monophyly, paraphyly, and natural kinds. Biology and Philosophy, 2005
↑ Willi Hennig
↑ abcd AMORIM, Dalton de Sousa. Fundamentos de Sistemática Filogenética (1º edição), Editora Holos, 2002
↑ ab Phylogenetic systematics. Urbana: University of Illinois Press, 1966
↑ POUGH, JANIS, HEISER. A Vida dos Vertebrados (4ª Edição),Editora Atheneu São Paulo, 2008
↑ Rosen; Nelson & Patterson. apud Hennig, 1979
Ligações externas |
- La Sociedad Willi Hennig
- Tree of Life Web Project
- Filogenetica.org: Plataforma de divulgación para la comunidad filogenética en Español


